Фрейд З. К пониманию афазий. Критическое исследование (1891)
[Перейти к странице сносок и примечаний]
V. [Глава пятая]
В одной из предыдущих глав [I] мы слышали, что теория речевого процесса Вернике в основе своей предусматривает особую роль центров в коре головного мозга, и [II] что клиника речевых нарушений не оправдывает вполне определенных ожиданий, которые вытекают из принятия такого положения. Это должно побудить нас внимательнее рассмотреть саму теорию.
Как и Вернике мы должны себе представить, что в коре больших полушарий головного мозга имеются определенные (точнее не ограничиваемые) зоны (зона Брока, зона Вернике), в нервных клетках которых содержатся каким-то образом представления, которыми оперирует речевая функция. Эти представления являются остатками впечатлений, которые прошли по пути зрительных и слуховых нервов или, как в случае движений языка (артикуляционных представлений) возникли на основе иннервационного чувства или восприятия осуществленного движения. Они располагаются вместе в коре головного мозга, в зависимости от происхождения из одного из этих источников. Таким образом, одна зона содержит все звуковые образы слов, другая все двигательные образы слов и т. д.[133]
Функция связи этих дистинктных центров возложена на белые волокнистые массы (ассоциативные пучки); и между центрами находится «незанятая зона» коры головного мозга, по выражению Мейнерта «функциональные пропуски».[134]
Вместе с последним определением мы покидаем путь развития мыслей Вернике и дополним его концепцию деталями из учения Мейнерта. Вернике, который не пропускал ни одной возможности сообщить, что его теория афазии представляет собой только применение идущего намного дальше учения Мейнерта, изначально предпочитал именно касательно речевых центров несколько отклоняющееся воззрение. В работе об афазическом симптомокомплексе (1874) ему все еще видится вся первая архаичная извилина вокруг Fissura Sylvii в роли речевого центра. В его учебнике болезней мозга (1887) напротив речевые центры представляются частями первой лобной и первой височной извилины. (Рис.7)
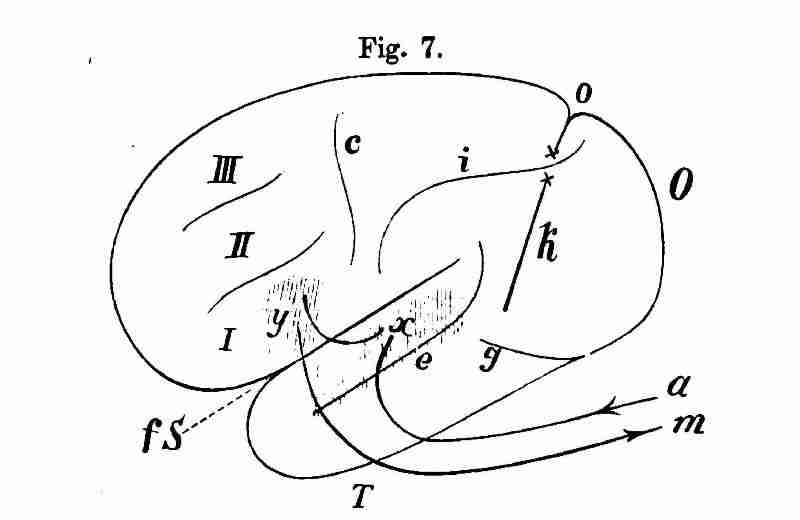
Рис. 7
В соответствии с Вернике, Учебник болезней мозга, Т.I, стр. 206. Рис.XX. Схема кортикального речевого механизма. F – лобное окончание[135], О – затылочное, Т – височное окончание левой гемисферы, fS Fissura sylvii. с – центральная извилина, g – передняя [136]окципитальная извилина, i – интерпариетальная извилина, k – передняя окципитальная извилина, о – парието-окципитальная извилина, е – параллельная извилина, I – III с первой по третью лобные извилины, x x — переходные извилины, x – сенсорный речевой центр, y – моторный речевой центр, xy — ассоциативный путь между двумя центрами, ax – путь акустики, ym – путь к речевой мускулатуре.
Мне кажется целесообразным сказать здесь несколько слов о научной концепции Мейнерта, о его представлении о мозговом устройстве и предназначении. Мое изложение этого учения, как и приводимые здесь же возражения представляют собой только бегло очерченные наброски, которые не могут соответствовать высокой значимости самого предмета. Однако, другой способ признания заслуг выходит далеко за рамки этой работы, в которой речь должна идти только о понимании речевых нарушений. Так как это понимание не может быть совсем независимо от общей трактовки деятельности мозга, я все же вынужден, по меньшей мере, затронуть вопрос о значении большого мозга.
Учение Мейнерта о строении головного мозга заслуживает наименование «кортико-центрического». В им же самим осуществленном комментарии анатомических взаимосвязей Мейнерт говорил, что кора головного мозга в силу поверхностного своего расположения предназначена для выполнения защитной, ограждающей функции и для принятия информации от всех органов чувств.[137] В дальнейшем она будет приравнена Мейнертом к сложной протоплазматической сущности, которая покрывает некое тело, чьи составляющие части она хочет ассимилировать, в то время как она сама преобразуется в полую изогнутую полусферу.[138] Весь прочий мозг представляется дополнением и вспомогательным органом коры больших полушарий головного мозга, и все человеческое тело является оснащением ее чувствительных нитей и улавливающих рук, которые предоставляют ей возможность принимать в себя образ внешнего мира и действовать согласно этому образу.[139]
[По Мейнерту] к коре головного мозга ведут или исходят из нее все волокнистые системы мозга. Все серые массы представляют собой лишь прерывающие течение волокнистых масс образования на их пути к большому мозгу. Спинной мозг исходит из коры большого головного мозга двойным началом, которое обнаруживается поперечным разрезом в области мозговой ножки. Так называемая ступня ножки содержит пути, которые передают двигательные импульсы коры головного мозга на периферию, а также пути, которые обусловливают принятие впечатлений от органов чувств в кору головного мозга. Она (ступня ножки) производит, таким образом, проекцию тела, и ставит его в непосредственную зависимость от структур коры головного мозга.[140] Так называемая крыша (Haube) мозговой ножки напротив сообщает коре головного мозга знание о рефлекторных процессах в спинном мозге и мозговом стволе и тем самым дает первый толчок собственным двигательным импульсам. Серые массы мозгового столба как четверохолмие и зрительный холм, которые имеют одновременно связь с нервными путями спинного мозга и обширными чувствительными поверхностями, представляют собой составные части того рефлекторного аппарата, который через крышу соединен с корой головного мозга. Или же они (как чечевичные зерна полосатого холма) в виде ганглий (нервных узлов) стопы ножки головного мозга прерывают прямой путь большого мозга. Специальный моторный канал, который подчиняет мускулатуру тела влиянию коры головного мозга, проходит от коры к периферии тремя отрезками (частями проективной системы), которые разделены двумя серыми узлами (чечевичное ядро- хвостовое ядро) и серой субстанцией передней роговицы. На мосту основной канал, кроме того, посредством серой субстанции, мостовых ядер и пучка волокон «пращевидной формы»[141] соединяется с малым мозгом, который иначе, согласно системе мозга Мейнерта, оставляется в стороне.Как формируется изображение тела в мозговой коре, которая, таким образом, связана с периферией? Мейнерт называет это изображение «проекцией»[142], и некоторые из его замечаний позволяют сделать вывод, что он и в самом деле предполагает проекцию, то есть изображение точка за точкой, объекта в мозговой коре. К примеру, в этом смысле столь частое сравнение мозговой коры с сетчаткой глаза[143], означает некий конечный орган, нервная субстанция которого многими авторами обозначается как «выдвинутая вперед часть серого вещества коры», в то время как по своей морфологии она должна была бы соответствовать части вещества спинного мозга. В пользу того, что термин «проекция» следует понимать в собственном смысле этого слова, дальше свидетельствуют несколько замечаний, такие как: «почти невероятным кажется предположение, что различные мускульные массы и поверхность кожи, железы и вещество выдающегося вперед пучка мозговой ножки так сильно рассеиваются для того только, чтобы потом быть представленными путем проекции на всей поверхности коры»[144], или: «ее (ножки) поперечный разрез охватывает (…) одновременно весь организм, который иначе был бы слеп и неспособен обонять»[145]. Все же дальнейшие разработки учения Мейнерта так сильно противоречат этой гипотезе, что я не вижу смысла здесь ее оспаривать.
Напротив, вряд ли можно ошибиться, если предположить, что Мунк[146] и другие исследователи, которые придерживались позиций Мейнерта, защищали более или менее ясно понимаемую идею совершенного и топографически похожего изображения тела в коре больших полушарий головного мозга.
Я могу лишь обратить внимание на то, что новейшие достижения анатомии головного мозга подтвердили в существенных аспектах понимание строения головного мозга Мейнерта и тем самым поставили под сомнение роль, приписываемую им коре головного мозга. Эти корректуры были связаны с происхождением наиболее значимого и лучше всего известного пути от коры головного мозга к мускулатуре тела. Сначала рухнуло понимание полосатого холма как одного из нервных узлов (ганглий) прерывающих моторный путь. Клиницисты, в первую очередь Шарко, показали, что его (моторного пути) повреждение влияет на подвижность только благодаря соседству с так называемой внутренней капсулой, и также не может вызвать паралич. Потом Вернике[147] указал на то, что эта так называемая ганглия мозговой ножки лишена сколько-нибудь многочисленных связей с корой головного мозга. Первый интернодиум оказался тем самым вырван из течения проекционного пути Мейнерта.
Изучение не одновременного пограничного образования (Markscheidenbildung) подтвердило этот факт и пробило новую брешь в понимании строения головного мозга Мейнерта. Флехсиг [1883] смог показать, что моторный путь от коры большого головного мозга к мускулатуре на самом деле не прерывается серым корковым веществом и проходит через внутреннюю капсулу в мозговую ножку, и что он (путь) на мосту ни в коем случае не имеет никакой связи с малым мозгом.
Пирамидный путь сейчас считается прямой связью между серым веществом передней роговицы (Vorderhorngrau) и серым веществом коры большого головного мозга; от утверждаемого Мейнертом вплетения малого мозга в моторный путь тем самым пришлось отказаться. Из больших субкортикальных масс только таламус оптикус в известной мере относится к коре большого головного мозга, но и он оказывается нежизнеспособным при врожденном дефекте тканей большого головного мозга; в то время как полосатый холм при дефекте большого мозга остается неповрежденным, но атрофируется при врожденной недостаточности малого мозга.[148] Большая часть субстанции головного мозга: полосатый холм-мозг противопоставляется, таким образом, большому головному мозгу как орган с неизвестной функциональностью, достаточно независимый от него по истории развития и функциональности. Объяснение Мейнертом двух этажей мозговой ножки тем самым больше не выдерживает критики, но, впрочем, до сих пор не заменено никаким другим. Если же говорить о двойном начале спинного мозга, то здесь, вероятно, имеется в виду начало большого мозга\таламуса и полосатого холма\малого мозга. Общее строение головного мозга указывает на наличие одной вершины в двух центральных аппаратах, из которых кора большого мозга представляется более поздним по возникновению, в то время как более древний аппарат ганглии переднего мозга, как кажется, сохранил часть своих функций. Также и другая важная составная часть учения Мейнерта, принятие положения о двукратном сенсибельном пути, одном прямом и одном рефлекторном, по всей вероятности подтверждена не будет. Наш опыт до сего дня учит нас, что ни один из рядов волокон не приводит к другой части мозга, не связавшись со спинным мозгом или аналогичным ему образованием, и что рефлекторные пути повсюду непосредственно исходят от сенсибельных.
Доминирующее положение коры большого головного мозга кажется, таким образом, поколебленным, и возникает необходимость перенести некоторые из считавшихся ранее субкортикальными процессов непосредственно в кору головного мозга.[149] Но тут возникает еще один вопрос, на который следует ответить, каким образом тело отображается в коре большого мозга. Как я полагаю скоро должно выясниться, что предположение о проекции тела в коре головного мозга в собственном смысле слова, под которым понимается совершенное и топографически похожее изображение, можно отклонить.
Я отталкиваюсь при этом от того феномена, на который Хенле обратил внимание еще в 1871 году при рассмотрении этого предмета, — редукции волокон серыми массами. Если сравнить количество волокон, поступающих в спинной мозг с тем, которое имеется в белом ответвлении пути, предназначенном для связи с высшими отделами головного мозга, то, очевидно, что количество последних составляет лишь небольшую часть первых. По подсчетам Штиллинга в одном из случаев 807738 волокон нервных корней соответствовали только 365814 волокнам верхней части шейного отдела спинного мозга в поперечном сечении.[150]Отношения спинного мозга к телу другого рода, нежели отношения серых масс верхнего отдела (к периферии тела). В самом спинном мозге (как и в аналогичных ему серых субстанциях) отношения к периферии представляют собой точную проекцию; каждое иннервационное единство на периферии может соответствовать одному фрагменту серой субстанции в спинном мозге, в крайнем случае, одному центральному элементу. Вследствие редукции проецирующих волокон серым веществом спинного мозга каждый элемент серой субстанции верхнего отдела не может более соответствовать одному периферическому единству, но должен соотносится со многими. Тот же самый закон распространяется и на кору большого головного мозга и здесь возникает необходимость обозначить два вида центральных репродукций разными именами. Если мы назовем репродукцию в серой субстанции спинного мозга проекцией, то будет подходящим назвать отображение в коре большого головного мозга репрезентацией и сказать, что периферия тела представлена в коре головного мозга не точка к точке, но выступает в менее детализированном виде отдельными волокнами.[151]Это до сих пор такое простое размышление получает дальнейшее развитие и другое направление благодаря следующим замечаниям:
Волокна верхней части шейного отдела спинного мозга в поперечном разрезе, в общем, предназначены не только для связи с корой головного мозга. Совсем не малая их часть (короткие пути) заканчиваются еще до окончания серого вещества полости между нервными центрами продолговатого мозга, другая часть попадает в малый мозг. Совершенно определенно мы можем сказать только о пирамидном пути, так как он предшествует шейному отделу спинного мозга, что он равно принадлежит к спинному мозгу и к серому веществу коры, и этот путь являет собой, конечно, значительно редуцированное продолжение волокон, которые от мускулатуры тела через передние корешки попадают в спинной мозг.
Редукция проецирующих волокон, с другой стороны не настолько велика, как это может показаться, после последнего замечания. Потому что, к примеру, вместо волокон задних путей, которые как таковые, конечно, не попадают в кору большого головного мозга, последняя (кора) принимает височные волокна, которые, после многократных прерываний в центрах задних путей, после серых масс продолговатого мозга и таламуса, наконец-то могут представлять задние пути в большом головном мозге. Неизвестно как соотносятся по численности височные волокна от волокон задних путей, вероятно, они (височные волокна) сильно отстают от них в этом плане.
Кроме того большой мозг получает волокна из малого мозга, в которых можно увидеть эквивалент начала малого спинного мозга, но, несмотря на все это, остается сомнительным, достигают ли коры головного мозга с периферии или кружным путем, в конце концов, столько же или больше волокон необходимых для осуществления проекции в спинном мозге.
Следует рассмотреть еще один аспект проблемы, который в учении Мейнерта проявляется не достаточно ясно. Для Мейнерта, который во всем потоке волокон видит, главным образом, важность самого факта корковых связей, волокна и волокнистые массы являются таковыми всегда, даже когда они уже миновали множественные серые субстанции. Его речевой оборот: «Волокно проходит серую субстанцию»[152], указывает на это. Возникает впечатление, как будто бы в волокне, на протяжении его долгого пути до коры ничего не меняется, даже когда оно вступает в многочисленные взаимосвязи.
Мы больше не можем придерживаться этого взгляда. Когда мы видим, как в процессе индивидуального развития происходит переход пограничного (прилегающего) образования (Markscheidenbildung) небольшими отрезками от одной серой субстанции к другой, как одному пути из серой субстанции соответствуют три и более ведущих дальше, нам уже кажется, что не пучки волокон, а именно серые субстанции являются отдельными органами мозга.[153] Если мы проследим сенсибельный (центростремительный) путь, насколько он нам известен, и признаем в качестве наиважнейшей его характеристики частое исчезновение в серых субстанциях и дальнейшее разветвление благодаря им[154], мы должны будем принять мысль о том, что волокно на пути к коре большого головного мозга меняется по своей функциональной значимости после каждого нового появления из серой субстанции.
Возьмем один из понятных примеров: волокно N. Opticus проводит одно из впечатлений глазной сетчатки к переднему четверохолмию; оканчивается в нем[155], и на его месте из серого вещества ганглии (нервного узла) исходит другое волокно к затылочному участку коры. В субстанции четверохолмия осуществилась связь оптического впечатления с двигательным восприятием глазного мускула; представляется очень вероятным, что новое волокно между четверохолмием и затылочной частью коры больше не проводит только впечатление глазной сетчатки, но доставляет в затылочную часть коры одно или несколько таких впечатлений, связанных с двигательным восприятием. Еще сложнее и значительнее должна быть структура изменения волокна применительно к проводящей системе кожной и мускульной сенсибельности; мы еще не имеем представления о том, какие элементы вступают во взаимодействие, чтобы сформировать новое содержание проводимого дальше возбуждения.
Здесь для нас принципиальным является то, что после прохождения через серое вещество волокна, достигающие коры головного мозга все еще имеют отношение к телесной периферии, но больше не могут дать топически верное отображение последней. Они содержат в себе телесную периферию, как – чтобы позаимствовать пример из актуального для нас сейчас предмета – какое-нибудь стихотворение содержит в себе алфавит, в измененном порядке, служащий совершенно другим целям, во всем многообразии связей различных топических элементов, одни из которых могут быть представлены многократно, другие же не проявиться вовсе.[156] Если бы можно было шаг за шагом проследить эту реорганизацию, которая осуществляется, начиная со спинальной проекции и до коры большого головного мозга, вероятно, обнаружилось бы, что ее принцип чисто функциональный и что топические моменты принимаются во внимание лишь в той мере
в какой они совпадают с требованиями функции. Так как ничто не свидетельствует о том, что в коре головного мозга происходит обратное изменение порядка, чтобы удовлетворить требованиям совершенной проекции, мы можем предположить, что телесная периферия в высших сферах мозга и в мозговой коре представлена больше не топически, но лишь функционально согласованно. Эксперимент на животных должен скрывать этот факт, хотя он может сообщить не что иное, как топическое отношение. Однако, я полагаю, что тот, кто всерьез ищет центр М. extensor pollucis longus, M. rectus internus oculi, или зону чувствительности определенного участка кожи в коре головного мозга, не признает также функцию этой зоны как совокупность условий, обеспечивающих существование этой функции.[157]
После такого отступления от темы мы[158] вновь возвращаемся к пониманию афазии и напомним себе (стр.21, 45), что на почве учения Мейнерта возникло предположение о речевом аппарате, состоящем из дистинктных центров коры, в клетках которой содержатся представления слов, чьи центры разделены нефункциональной областью коры и соединены белыми волокнами (ассоциативными связками). Можно с самого начала поднять вопрос о корректности и допустимости предположения подобного рода, которое заключает представления в клетки. Я нахожу его некорректным.[159]
В противоположность тенденции эпохи ранней медицины локализовать все душевные процессы, в том виде как они разграничены психологией языкового употребления, в определенных зонах мозга, учение Вернике, несомненно, представляло собой огромный шаг вперед. Он утверждал, что локализовать, а именно в центральных окончаниях периферических нервов, которые принимают впечатления,[160] позволительно только простые психические элементы, отдельные чувственные представления. Не была ли, однако, по сути, сделана та же самая принципиальная ошибка, попытка вместить сложное понятие в узкие рамки, локализовать как единый психический элемент всю душевную деятельность? Правомерно ли нервные волокна, которые на протяжении своего долгого пути были подчинены только физиологическим структурам и модификациям, в конце этого пути окунать в сферу психического и нагружать их там представлениями или образами памяти? Если «воля» и «интеллигентность» и тому подобное известны как психологически искусственно образованные слова, которым в физиологическом мире соответствуют очень сложные отношения, то о «простом чувственном представлении» с большой степенью уверенности можно сказать, что оно все же являет собой нечто отличное от такого искусственно образованного слова?
Последовательность физиологических процессов в нервной системе, по всей вероятности, не находится в отношениях причинности к психологическим процессам. Физиологические процессы не прекращаются одновременно с началом психических, во многом развитие физиологической цепи продолжается, с той только разницей, что каждому элементу последней (или отдельным звеньям), начиная с определенного момента, соответствует психический феномен. Психическое, тем самым, являет собой параллельный процесс физиологическому[161] («a dependent concomitant»).[162]
Я прекрасно понимаю, что не могу требовать от людей, чьи взгляды я сейчас оспариваю, чтобы они без возражений приняли столь резкий скачок и полную перемену курса научных воззрений.[163] Они же, очевидно, предполагают, что — физиологическая составляющая – модификация нервных волокон, при чувственном возбуждении становится причиной другого изменения в центральной нервной клетке, которая и становится физиологическим коррелятом «представления». Так как о представлении она (клетка) может сообщить гораздо больше, чем о физиологически еще неохарактеризованных неизвестных модификациях, последние описываются использованием эллиптического выражения: якобы в нервной клетке локализовано представление. Одно только это замещение тотчас же и приводит к смешению обоих понятий, которые между собой по сути дела имеют мало что общего. В психологии простое представление для нас являет собой нечто элементарное, что мы без труда можем отделить от его связей с другими представлениями. Таким образом, мы приходим к предположению, что и его (представления) физиологический коррелят, модификация, им обусловленная, исходящая из оканчивающихся в центре волокон, также есть что-то простое, сводимое к локализации в одном пункте. Такое перенесение, разумеется, абсолютно неправомерно[164]; свойства этой модификации должны определяться сами по себе, вне зависимости от ее психологического эквивалента.[165]
Однако, что является физиологическим коррелятом простому или для нее (психологии) возвращающемуся представлению? Очевидно, что это нечто не константное, но имеющее в основе своей природы динамический процесс. Этот процесс в состоянии вынести локализацию, он исходит из особой зоны коры головного мозга и распространяется оттуда по всей коре или вдоль особых путей.[166] Когда этот процесс окончен, он оставляет за собой в зоне коры головного мозга, подвергшейся его воздействию, модификацию, возможность воспоминания.[167] Все же очень сомнительно, что эта модификация в то же время находит свое соответствие в психической сфере; наше сознание не регистрирует со стороны психического ничего, позволяющего оправдать введение наименования «латентного воспоминания».
Но, как только то же самое состояние коры воспроизводится снова, психическое возникает заново в виде воспоминания. Мы, конечно, не имеем ни малейшего представления о том, как можно привести животную субстанцию в состояние, когда она способна производить и различать столь многофактурные модификации. Но то, что она это действительно может делать, доказывает пример сперматозоидов, в которых заложена многофактурная и детальнейшая модификация дальнейшего развития.
Можно ли у физиологического коррелята ощущения найти какую-то часть «восприятия», отличную от «ассоциации»? Очевидно, нет. «Восприятие» и «ассоциация» представляют собой два наименования, которые мы нагружаем различными воззрениями касательно одного и того же процесса. Но нам известно, что оба обозначения абстрагированы от единого и нерасчленимого процесса. Мы не можем получить какое-либо восприятие, не ассоциируя его при этом; и хотя на уровне понятий мы способны их так четко разделить, в действительности они принадлежат единому процессу, который, начинаясь в одной зоне, диффундирует по всей коре. Локализация физиологического коррелята являет собой, тем самым, по отношению к представлению и ассоциации демонстрацию того факта, что локализация представления означает не что иное, как локализацию его коррелята. Таким образом, нам следует отказаться от того, чтобы относить представление к одному участку коры головного мозга, ассоциацию же переносить в другой. Оба исходят в большинстве своем из одной зоны, и ни в какой зоне не остаются постоянно. С этим отказом признавать какую-то особую локализацию процессов образования и ассоциирования представлений, для нас исчезает основной повод проводить границу между центрами и проводящими путями языка. В любой зоне, служащей реализации речевой функции, будут предполагаться подобные же функциональные процессы, и нам совсем не нужно, привлекать белые волокнистые массы и поручать им осуществление ассоциативной связи находящихся в коре представлений. Мы даже располагаем данными судебно медицинского вскрытия, которые доказывают, что ассоциация представлений происходит по путям, располагающимся в самой коре. Я опять имею ввиду случай Хойбнера, который уже позволил нам сделать одно важное заключение.(срав.стр.32).
Больной Хойбнера демонстрировал ту форму речевого нарушения, которую Лихтхайм обозначал как транскортикальную сенсорную афазию и обосновывал прерыванием путей от сенсорного центра языка к ассоциации понятий.[168] Было бы логично ожидать в свете вышеупомянутой теории заболевание внутренней ткани ниже сенсорного центра. Вместо этого обнаруживается поверхностное размягчение коры, которое само (даже согласно функции) отделяет неповрежденный сенсорный центр от большинства его связей в коре за пределами речевой зоны. Хойбнер не преминул подчеркнуть важность этого открытия, а Пик [169]вывел из него то же самое заключение, что и мы, а именно вероятность прохождения ассоциативных путей речи по самой коре. Впрочем, нам нет необходимости оспаривать тот факт, что ассоциативные пучки, которые проходят ниже в коре также могут служить выполнению этой функции.
Наше представление о речевом аппарате должно будет претерпеть существенное изменение, когда мы рассмотрим еще и третье утверждение учения Мейнерта – Вернике, собственно то, которое разделяет функциональные речевые центры «нефункциональными пропусками».[170] Сначала могло бы показаться, что это положение, как взятое из непосредственных данных патологической анатомии, сомнению не подлежит.
Но если задуматься о способе, с помощью которого на основании данных судебно медицинского вскрытия был сделан вывод о существовании дистинктных центров, можно заметить, что патологическая анатомия не в состоянии ответить на этот вопрос. К примеру, можно бросить взгляд на таблицу, в которой Наунин[171] фиксировал величину и расположение повреждений в семидесяти одном случае речевых нарушений. Там, где повреждения очень плотно накладывались друг на друга, мы предположили наличие центров речи. Эта дефиниция произведена согласно зонам, которые должны быть непременно задействованы для выполнения определенной речевой функции. Но при этом могут быть задействованы и другие зоны коры, одновременно служащие выполнению этой же функции, разрушение которых, однако, легче переносится речевой функцией. Если такие корковые зоны существуют, мы не сможем найти их, изучая таблицу Наунина. Вполне возможно, что вызванное травмой речевое нарушение является вариантом такого удаленного воздействия[172] других зон, достигшего определенного центра. Не исключено и то, что реже подвергающиеся травмированию (согласно таблице) зоны, также являются «речевыми центрами», пусть и менее необходимыми или константными.
Давайте все-таки вернемся к вопросу о том, какое влияние приписывается авторами нефункциональной области коры, располагающейся в непосредственной близости от речевых центров и между ними.
Мейнерт говорит об этом совершенно определенно (Психиатрия (1884), стр.140): «Отсюда, конечно, следует, что в течении физиологического процесса оккупации коры головного мозга воспоминаниями, происходит увеличение нагрузки на клетки коры, которые являются основой дальнейшего развития круга детских представлений, осуществляемого через обогащение образов памяти. Очень вероятно, что памяти, как основе всех интеллектуальных достижений, тоже положен предел, в виде «ограничения принимающей способности клеток коры головного мозга». Последнее высказывание можно отлично интерпретировать в том смысле, что не только детское развитие, но и приобретение позднейших знаний (к примеру, изучение какого-нибудь нового языка), строится на почве процесса оккупации незанятых до того мест в коре головного мозга,[173] примерно как расширение города осуществляется путем вынесения поселений на какое-то расстояние за городские стены.
В одном из более ранних замечаний этим, соседствующим с центрами, но еще незанятым областям, Мейнертом присваивалась способность принятия на себя функций разрушенных посредством экспериментов или иным образом центров[174]. Это воззрение опиралось на опыты Мунка, одного из тех исследователей, чьи предположения корнями уходили в почву учения Мейнерта. Итак, сейчас мы знаем, с каких позиций было сделано предположение о существовании «функциональных пропусков» в коре головного мозга, и, исходя из этого, можем сделать попытку оценить пригодность подобного понимания речевых нарушений.
Мы обнаружим при этом нечто, в корне противоположное тому, что соответствовало бы ожиданиям после принятия такого базового предположения. Речевая функция демонстрирует отличнейшие примеры новоприобретений. Уже обучение чтению и письму дает нам пример таковых в оппозиции к первоначальной речевой деятельности. Это новоприобретение должно инициировать повреждение через образование новой локализации, так как в этом случае наблюдается привнесение новых чувствительных элементов (оптических и сенсо-моторных).
Все дальнейшие новоприобретения речевой функции – учусь ли я понимать и говорить на иностранных языках, осваиваю ли наряду с первым буквенным алфавитом греческий или иудейский, изучаю ли наряду с моей курсивно-шрифтовой стенографией другое письмо – все эти достижения (по количеству применяемых для их осуществления образных воспоминаний они могут с полной очевидностью многократно превосходить первоначальный язык) локализуются на том же самом месте, которое нам известно как центр первичного изучения языка. Никогда ведь не происходит такого, чтобы органическое повреждение обусловило бы нарушение функции родного языка, обойдя своим вниманием языки, освоенные позднее.[175]Если бы у какого-нибудь немца, который понимает также и французский, французские словесно звуковые образы были бы локализованы отдельно от немецких, рано или поздно должно было бы случиться так, что вследствие размягчения очага локализации, немец не понимал бы больше немецкого, но понимал бы французский. На самом же деле для всех без исключения языковых функций характерно как раз обратное положение вещей.[176] Когда я привожу соответствующие (к сожалению, учитывая ваш интерес, не слишком многочисленные) примеры, я обнаруживаю два момента, которые могут обусловить появление речевых нарушений у субъектов, говорящих на многих языках: 1. Влияние давности изучения языка, 2. (Частота) его использования.[177] Как правило, оба момента действуют одновременно. Там же, где встречается противоречие, более ранняя по времени приобретения языковая способность удивительным образом может превзойти по длительности сохранения даже чаще применяемую. Однако, никогда не встречается зависимость, которая должна была бы объясняться различной локализацией, а не влиянием двух вышеозначенных функциональных моментов. Дело, очевидно, обстоит так, что языковые ассоциации, с которыми работает наш языковой потенциал, способны к сверхассоциированию, процессу, который мы ясно ощущаем, в то время, когда производство новых ассоциаций дается нам с трудом. Ясно также, что процесс сверхассоциирования при любом расположении повреждений оказывается нарушен раньше, чем первичное ассоциирование.[178]
То, каким образом редко, но интенсивно производимая модификация речевого аппарата[179] может пережить повреждение, вряд ли можно продемонстрировать более впечатляюще, чем на следующем примере, который я заимствую у Хуглингс Джексона. Этот исследователь, к чьим воззрениям я постоянно возвращался почти во всех предшествующих замечаниях, чтобы с их помощью оспорить локализаторную теорию речевых нарушений, приводит совсем не редкий случай, когда моторно-афазические больные кроме «да» и «нет» имели в своем распоряжении и другие остатки языка, которые вообще то должны были бы соответствовать высокоразвитым языковым функциям.[180] Эти языковые остатки нередко представлены крепкими ругательствами (Sacre nom de dieu, Goddam etc.)[181], и Хуглингс Джексон заключает, что последние принадлежат к здоровому в эмоциональном плане, и интеллектуально неиспорченному языку. В других случаях этот языковой остаток представляет собой не проклятье, но слово или часть речи с очень узким значением, и следовало бы удивиться тому, что именно эти клетки или эти воспоминания столь счастливо избегли всеобщего разрушения. Некоторые из этих случаев допускают вполне убедительное истолкование. К примеру, человек, который мог сказать только: «I want protection»[182] (что-то вроде: Я прошу, на помощь), был обязан своей афазией драке, во время которой, после удара по голове, упал без сознания. У другого имелся в высшей степени примечательный речевой остаток «List complete»[183]. Он был писателем, которого настигла болезнь после того, как он, работая по очень интенсивному графику, закончил каталог. Такие примеры позволяют допустить предположение, что эти речевые остатки представляют собой слова, которые речевой аппарат образовал перед заболеванием, возможно даже предчувствуя его.[184] Я стремлюсь объяснить сохранение этой последней модификации внутренней интенсивностью переживания ситуации, которой сопровождалось ее возникновение. Я припоминаю[185] те два случая, когда я полагал, что моей жизни угрожает опасность. Это ощущение оба раза возникало совершенно внезапно.
В обоих случаях я подумал про себя: «Всё, сейчас тебе конец», и, в то время, когда промелькнула моя внутренняя речь, обычно сопровождающаяся лишь неясными звуковыми образами и почти неразличимыми ощущениями на губах, в момент опасности я услышал эти слова так, как будто кто-то прокричал мне их в ухо и одновременно увидел их как будто напечатанными на фланирующем листке бумаги.
Таким образом, мы отказываемся от предположения, что речевой аппарат состоит из обособленных центров, которые разделены нефункциональными областями коры. Далее мы отвергаем также и то допущение, что в определенных местах коры головного мозга, называемых центрами, складируются представления (воспоминания), которыми оперирует речь, в то время как ассоциация осуществляется исключительно за счет белых волокнистых масс, расположенных под корой. Теперь единственное, что нам остается – это высказать предположение, что речевая зона коры представляет собой связную взаимозависимость всех областей коры головного мозга, внутри которой ассоциации и перенесения, являющиеся основой речевой функции, существуют во всей, подробнее не доступной пониманию, сложности.[186]
Исходя из этого предположения, как мы будем объяснять существование речевых центров, прежде всего зон Брока и Вернике, наличие которых подтверждается патологией связанных с ними нарушений? Здесь можно обратить свой взор на выпуклую поверхность левой гемисферы.[187] Так называемые центры речи выявляют определенные, требующие пояснения, отношения, суть которых можно обнаружить, опираясь на нашу точку зрения.
Они располагаются достаточно далеко друг от друга. Если следовать Наунину, в задней части первой височной извилины, в задней части третьей лобной извилины, внизу теменной дольки, где Gyrus angularis переходит в затылочную долю[188]. Расположение четвертого центра двигательных образов письма еще установлено не достаточно точно (задняя часть средней лобной извилины?). Они настолько удалены друг от друга, что заключают между собой обширную область коры (остров с покрывающими его фрагментами извилины), повреждение которой всегда связано с нарушениями речи. И, несмотря на то, что протяженность этой области, в результате сопоставления обнаруженных при афазии повреждений, точно не ограничена, все же можно утверждать, что она образует внешние участки предполагаемой нами речевой зоны. Речевое нарушение, исходя от центров, проходит внутрь (где-то посередине дуги гемисферы), в то время как с внешней стороны участков расположены корковые зоны другого предназначения.
Таким образом, «центры» являются углами речевого поля, и далее, следует обратить внимание, на те области, в которые эти центры упираются снаружи. Зона Брока расположена в непосредственной близости моторных центров для утолщенных нервов (Bulbarnerven); зона Вернике находится в области, содержащей акустические окончания, точное местоположение которых неизвестно, визуальный центр упирается в зоны затылочной доли, в которых мы привыкли искать окончания Nervus opticus. Такая организация, не имеющая смысла, если исходить из теории центров, нами объясняется следующим образом:
Ассоциативная область языка, в которую входят оптические, акустические и моторные (или кинестетические) элементы, располагается между корковыми полями этих чувствительных нервов и соответствующими моторными корковыми полями. Если в таком ассоциативном поле мы представим себе сдвигающееся повреждение, то оно будет производить тем больший эффект (при равной величине), чем ближе оно будет придвигаться к одному из корковых полей, чем более близко к периферии оно будет располагаться в речевом участке. Если оно затронет непосредственно одно из этих корковых полей и, таким образом, перережет один из каналов ассоциативной области, в речевом механизме будет отсутствовать оптический, акустический и т.д. элемент, потому что каждый ассоциативный импульс этого рода исходит из упомянутого коркового поля. Если же передвинуть повреждение дальше внутрь ассоциативного поля, эффект будет менее выраженным, и ни в коем случае не сможет уничтожить все ассоциативные возможности этого вида. Таким образом, части речевого поля, упирающиеся в корковые поля оптических, акустических и моторных мозговых нервов обретают то значение, которое выявляет патология, и которое привело к их выдвижению на первый план как центров речи. Это значение имеет смысл только в рамках патологии, но не распространяется на физиологию речевого аппарата, поэтому, нельзя утверждать, что в них происходят другие, или более значимые процессы, чем в тех частях речевого поля, повреждения которых легче переносятся. Эта гипотеза является непосредственным следствием отказа отделить процесс построения представлений от ассоциирования и локализовать эти процессы в разных зонах.
Вернике близко подошел к такой точке зрения, когда он в своих последних высказываниях на данную тему усомнился в правильности предположения, согласно которому чтение выделялось в особый центр внутри оптических корковых окончаний, а письмо относилось к моторному, так называемому ручному региону (Armregion). Его размышления, однако, не имели принципиального характера, и учитывали только анатомическую поправку, согласно которой важные для речи оптические и хейромоторные (cheiromotorischen) воспоминания разрозненно существовали внутри других образов памяти, той же природы. Хойбнер, напротив, уделив внимание сообщенному им случаю, вплотную приблизился к сомнительному вопросу касательно речи, который аналогичен нашему[189]: «Или, может быть, вовсе не существует корковых полей, с которыми связана визуальная, акустическая нечувствительность, паралич? Возможно, симптом такого состояния возникает в гораздо большей степени за счет того, что корковые поля, непосредственно служащие для осуществления данных функций (…), изолируются от остальной коры головного мозга соседним очагом размягчения?»
Все же нам следуем ответить еще на два возражения, которые могут быть направлены против нашего понимания центров. 1. Если разрушение фрагмента речевой зоны, которая непосредственно примыкает к корковому полю (оптики, акустики, руки, языка и т.д.), имеет манифестируемые последствия для речевой функции только в силу того, что прервана связь с оптическими, акустическими и другими ассоциативными импульсами, тогда ведь разрушение самого этого поля должно обусловливать те же последствия для речи. Однако, это прямо противоречило бы нашему опыту, который показывает наличие локального симптома при подобных повреждениях без нарушения речи. Это первое возражение нейтрализовать легко, если предположить, что все другие корковые поля представлены в удвоенном варианте, а ассоциативное поле речи структурируется только в одной гемисфере. К примеру, разрушение оптического поля коры не помешает реализации визуального импульса речи (чтения), потому что речевое поле при этом сохраняет свои связи (на этот раз осуществляемые белыми перекрестными волокнами) с оптическим полем коры на другой стороне. Если же повреждение сдвигается к границе оптического коркового поля, возникает алексия, потому что разрывается связь не только с оптическим корковым полем той же стороны, но и с перекрестным оптическим корковым полем противоположной. Таким образом, нам следует ввести сюда предположение, что видимость наличия центров возникает благодаря тому, что перекрестные связи корковых полей другой гемисферы сосредотачиваются в тех же самых местах, а именно на периферии речевого поля, где проходит связь с корковым полем той же стороны. Это звучит убедительно, так как для реализации речевой ассоциации двойное представительство оптических, акустических и других импульсов не имеет никакого физиологического значения.
Впрочем, это совсем не новое предположение, скорее позаимствованное из теории центров, которая утверждала наличие такой связи между речевым участком и двусторонними корковыми полями. Собственно говоря, анатомические отношения этой перекрестной ассоциации еще не установлены, и могут прояснить некоторые особенности в расположении и протяженности кажущихся центров, как и индивидуальный характер проявлений речевых нарушений.
- Можно было бы поинтересоваться, зачем так уж необходимо, оспаривать наличие каких бы то ни было речевых центров, если мы при этом вынуждены обсуждать корковые поля, то есть центры оптики, акустики и моторных речевых органов? На это можно возразить тем, что подобные рассуждения все равно повторились бы в отношении других так называемых моторных и чувствительных корковых центров, что существование даже более резко очерченных корковых полей невозможно оспорить в отношении других функций, потому что последние могут быть охарактеризованы анатомическим фактом окончаний чувствительных нервов или предъявлением соответствующей части пирамидного пути в определенных областях коры головного мозга. Ассоциативному полю речи все же не хватает таких прямых отношений к периферии тела. Оно не располагает собственными сенсибельными и, с высокой степенью вероятности, также особыми моторными «проективными путями»[190]. [191]
[Конец пятой главы]
[Перейти к шестой главе]
[Перейти к странице сносок и примечаний]
























